Trwa ładowanie...
Money.pl
Zaloguj
Giełda
Notowania GPW
Spółki GPW
Porównanie spółek
Giełda na żywo
Giełdy światowe
Komunikaty ESPI
Surowce
Narzędzia giełdowe
Portfel
Waluty
Kursy walut
Kalkulator walutowy
Bitcoin
Forex
Depozyty
Dla przedsiębiorców
Jak założyć firmę
Faktoring dla firm
Poradniki
PKD
PKWiU
Mobility
Praca
Oferty pracy
Wynagrodzenia
Kalkulator PPK
Kalkulator wynagrodzeń
Biznes i Gospodarka
BizTech
Ekologia, OZE, ESG
Nieruchomości i budownictwo
Emerytury
Prawo
Kariera i rynek pracy
Finanse osobiste
Kalkulatory
Kalkulator urlopu macierzyńskiego
Kalkulator odsetkowy
Kalkulator zakupów na firmę
Kalkulator składek ZUS
Kalkulator wynagrodzenia za pracę w nadgodzinach
Kalkulator dat
Kalkulator wynagrodzenia chorobowego
Kalkulator ekwiwalentu za urlop wypoczynkowy
Kalkulator stażu pracy
Kalkulator kredytu hipotecznego
Kalkulator składek ZUS dla działalności gospodarczej
Strefa money
Biznes Klasa
Program Money
Nagroda Money
Pomysł Na Dom
Stan gospodarki
Wybory parlamentarne 2023
Obietnice wyborcze 2023
Money Insights
Wydarzenia
Impact
Kongres 590
Forum Ekonomiczne
Szczyt Klimatyczny Togetair
Zapytaj Notariusza
Krynica Forum 2022
Krynica Forum 2023
Rynek nieruchomości - INPON 2023
Wyższy poziom chmury
Cybersec Forum
Kongres ESG Polska Moc Biznesu
II Europejski Kongres Sportu I Turystyki
Podatki
Wiadomości podatkowe
Pit
Formularze podatkowe
Poradniki podatkowe
Znajdź Adres
Prawo
Wiadomości
Twoje finanse (REKLAMA)
Fundusze
Wiadomości
Profile funduszy
Wyszukiwarka fundusz
Stopa zwrotu z funduszy
Kalkulator prowizji
Kalkulator zysku
Archiwum Funduszy
Banki
Poradniki
Ubezpieczenia
Emerytury
Poradniki
Giełda
NARZĘDZIA
Notowania GPW
Spółki GPW
Porównanie spółek
Giełda na żywo
Giełdy światowe
Komunikaty ESPI
Surowce
Narzędzia giełdowe
Portfel
Waluty
NARZĘDZIA
Kursy walut
Kalkulator walutowy
Bitcoin
Forex
Depozyty
Dla przedsiębiorców
Jak założyć firmę
Faktoring dla firm
Poradniki
NARZĘDZIA
PKD
PKWiU
Mobility
Praca
NARZĘDZIA
Oferty pracy
Wynagrodzenia
Kalkulator PPK
Kalkulator wynagrodzeń
Biznes i Gospodarka
BizTech
Ekologia, OZE, ESG
Nieruchomości i budownictwo
Emerytury
Prawo
Kariera i rynek pracy
Finanse osobiste
Kalkulatory
NARZĘDZIA
Kalkulator urlopu macierzyńskiego
Kalkulator odsetkowy
Kalkulator zakupów na firmę
Kalkulator składek ZUS
Kalkulator wynagrodzenia za pracę w nadgodzinach
Kalkulator dat
Kalkulator wynagrodzenia chorobowego
Kalkulator ekwiwalentu za urlop wypoczynkowy
Kalkulator stażu pracy
Kalkulator kredytu hipotecznego
Kalkulator składek ZUS dla działalności gospodarczej
Strefa
Biznes Klasa
Program Money
Nagroda Money
Pomysł Na Dom
Stan gospodarki
Wybory parlamentarne 2023
Obietnice wyborcze 2023
Money Insights
Wydarzenia
Impact
Kongres 590
Forum Ekonomiczne
Szczyt Klimatyczny Togetair
Zapytaj Notariusza
Krynica Forum 2022
Krynica Forum 2023
Rynek nieruchomości - INPON 2023
Wyższy poziom chmury
Cybersec Forum
Kongres ESG Polska Moc Biznesu
II Europejski Kongres Sportu I Turystyki
Malwina Gadawa
Przedsiębiorcy odetchną? Tak będą kontrolowani. Znamy szczegóły
Karuzela kadrowa znów ruszyła. Komu przypadnie Pekao?
Polski węglowy gigant ma nowego prezesa
Fala zwolnień zalewa Polskę. Kiedy się zatrzyma? "Dwa sprzeczne trendy"
Bank Pekao podjął kluczową decyzję. Rada nadzorcza w nowym składzie
Kto będzie nowym prezesem ZUS? Ujawnili nazwisko. Nieoficjalne informacje
MAT. SPONSOROWANY
Pierwszy kredyt firmowy. Jak się do niego przygotować?
Money Magazyn
Czarna zemsta ajatollahów. Iran wciąż nie powiedział ostatniego słowa
Przemysław Ciszak
"Zarabiałem 3,8 zł za godzinę". Dziś ma firmę z przychodem 200 mln zł
Łukasz Kijek
Money Insights: Rząd ma problem wart 250 mld zł [OPINIA]
Marcin Mrowiec
Gospodarka
Ukraińcy z Polski na front? Jest podpis. Kijów może pójść krok dalej
Paweł Gospodarczyk
Pytają o inwestora w browarze Leżajsk. Boją się, że to firma "słup"
Ukraina w końcu dostanie wsparcie ze Stanów? "Przyszedł czas na działanie"
Dziura w finansach na wiele lat. MFW ma przykrą prognozę dla Polski
Świat wstrzymał oddech. Ruchy Iranu mogą rykoszetem uderzyć w portfele Polaków
"Z chmur na ziemię". Pełnomocnik rządu mówi, co się stanie z głośnym projektem PiS
Dodatkowe sankcje na Iran. Obejmą podmioty gospodarcze
Finanse
Ceny prądu. Wiadomo ile wyniesienie maksymalna stawka
MLG
W tej kategorii polski gigant nie ma sobie równych. Jest nowy ranking
Miliardy na wymianę okien i ocieplenie. Minister ogłosiła
Zbliża się ważny termin ws. 800 plus. Wielu rodziców nadal zwleka
Ceny prądu. Wiadomo już, o ile wzrosną rachunki
Bon turystyczny. Minister Nitras: nie ma powodu do wznawiania
Zmiany w podatku Belki. Budżet straci 1,5 mld zł
Notowania
GIEŁDA
WALUTY
SUROWCE
Biznes
Polskie firmy w Izraelu. Na pierwszy plan wysuwa się jeden problem
"Nie jesteśmy tylko liczbami". 650 ludzi trafi na bruk. Piszą do zarządu giganta
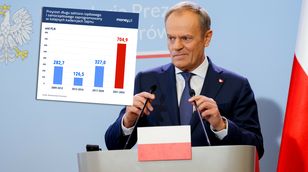
Urząd tłumaczy zatrudnienie Michała Tuska. Znamy zakres obowiązków syna premiera
Zamieszanie wokół spółek Skarbu Państwa. Co z obietnicą Donalda Tuska?
Paraliż jednego z największych lotnisk na świecie. Emirates odwołują wyloty z Dubaju
Rynki
Giełda
Waluty
Rekordowe ceny złota. Inwestorzy wiedzą swoje
Europa bliżej przełomowej decyzji. Inflacja w strefie euro i UE spada
Załamanie na rynku. Takich przewidywań cen złota jeszcze nie było
Kursy walut 17.04.2024. Środowy kurs funta, euro, dolara i franka szwajcarskiego
Ile kosztuje dolar? Kurs dolara do złotego PLN/USD 17.04.2024
Wideo
Biznes Klasa
Pomysł na dom
TYLKO W MONEY
Zaczynał na stadionie. Dziś ma 120 salonów i obraca milionami
Łukasz Kijek
Zarządza potężnym majątkiem polskiego miliardera. "Złoto się kończy"
Spytaliśmy młodych. Wiedzą, ile kosztują prąd, benzyna czy iPhone?
Rosja stawia wszystko na jedną kartę. "Polska musi zrobić trzy rzeczy"
Prawo
Rosja domaga się bankructwa PGNiG. Pozew z kosmosu
Paweł Gospodarczyk
Konfiskata aut za alkohol. Rząd robi pół kroku w tył. Jest projekt
Brexit dopiero teraz uderzy w Wielką Brytanię? Do wielkiej zmiany zostały dni
Radia apelują do KRRiT. Zaległości wynoszą ponad 66 mln zł
Energetyka
Zwrot na stacjach. Kierowcy nie widzieli tego od miesiąca
Chińskie panele i wiatraki już zalewają Europę. UE bije na alarm
Zmiany w programie Czyste Powietrze. "Niezadowolenie i niepewność"
Praca i kariera
Ludzie Macieja Laska przechodzą z CPK do PPL. Kolejna osoba zrezygnowała
Tak chcemy być nagradzani w pracy. Badanie mówi jasno
W polskich szkołach pojawią się żołnierze. Przeszkolą uczniów
Wybrane dla Ciebie
Najnowsze
Biznes
Finanse
Księgowość
Goesgreen
Zarobki